Journal of Multidisciplinary Dental Research
Volume: 4, Issue: 2, Pages: 64-70
Review Article
Archana Vaikom Krishnan1, Shashidara Raju2, Sudheendra U S3, Aparna H G4
1Senior Lecturer, Oral Pathology, Microbiology and Forensic Odontology, Coorg Institute of Dental Sciences, Virajpet, Karnataka, India
2Head of the Department, Oral Pathology, Microbiology and Forensic OdontologyCoorg Institute of Dental Sciences, Virajpet, Karnataka
339, Vyasa Sadana, Theobald Road, NAzarbad, Mysore, 570010, Karnataka, India.
4Oral Pathology, Microbiology and Forensic Odontology,Coorg Institute of Dental Sciences,Virajpet, Karnataka
Corresponding
Archana Vaikom Krishnan
Senior Lecturer, Oral Pathology, Microbiology and Forensic Odontology
Coorg Institute of Dental Sciences, K K Campus, Maggula Post, Virajpet- 571218, Karnataka, India
E-mail: [email protected]
Phone: +91-9791300362
Received Date:24 November 2018, Accepted Date:12 December 2018, Published Date:25 December 2018
Viral infections have the unique property of identifying themselves on histopathological slides by morphologically altering cells. Koilocytes are one such cytopathies that are a result of Human Papilloma Virus (HPV) infection. The cell has gained popularity in recent times over the increased incidence of HPV associated oral squamous cell carcinomas, yet little is discussed regarding koilocytosis and its significance. This review focusses on the basis of koilocytosis with special reference to the role of viral proteins in cell structure alteration with a note on prognostication.
Viral transformation:
Over the years the understanding of viral infections and subsequent influences on infected cells has expanded multifold from light microscopic justification with inclusion bodies, up to the stage of molecular understanding of tumour virus induced oncogenesis. Viruses tend to have one of three effects on the cells they infect- Damaging the cell, persisting within it in a state of latency or transforming it. Morphological alterations may occur in the form of a) cell surface membrane alterations giving rise to multinucleated cells as we see in measles (Warthin-Finkleday cells); b) Inclusion bodies: intra-cytoplasmic (eosinophilic Rhabdoviral inclusions of the neurons- Negri bodies) or intranuclear(herpes);c) Neooplastic transformation; and d) cell death.1, 2
Numerous tumor viruses show the property of transforming- both functionally and morphologically, cells that they infect. This process of transformation occurs when the cell in particular does not, for some reason, allow the viral particle to replicate within it. This leads to the integration of the viral DNA into the resisting cell causing mutation and ensuing transformation.1
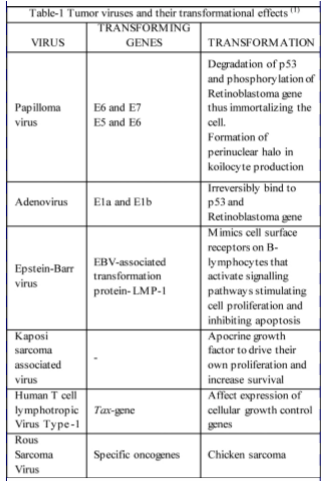
Most studies on virally induced cell transformation and oncogenesis were carried out using the Simian Virus-40 and Polyoma-viruses, which with their small nucleic acid size (5kb) serve as useful tools in elucidating mechanisms of the same. Through continued research it was demonstrated that the parts of the viral genome that induce cell lysis in infected cells were the same that effect cell transformation. The first expressed genes are the early genes that encode for viral DNA synthesis, stimulate host cell gene expression and enzyme production required for viral replication and the late region genes are expressed later and encode for synthesis of the structural components required for virion packaging. Table-1 highlights some important tumor viruses their transforming genes and their effects.1
HPV and its cytopathy:
A member of the Papovavirus family, the human papilloma viruses are a large family of oncogenic and non-oncogenic viral particles. They are double stranded DNA viruses that show selective tropism for and infect the squamous epithelia causing mostly papillomatous lesions. Long being implicated in the pathogenesis of cervical carcinomas, they have now for the past decade been extensively studied for their involvement in oropharyngeal carcinomas.
The life cycle of the HPV begins with its inoculation into wounded discontinuous epithelium, followed by infection of mitotically active basal cell layers. The virus then immortalizes the cell though its early genes E6 and E7 allowing the cell to remain mitotically active as it traverses to the top of the epithelium at the same time providing for its intra-nuclear replication. During its life span the virus morphologically alters the cells, through mechanism discussed later, that give rise to pathognomic cytopathic effects that allow for morphological diagnosis of HPV infections and related lesion.
One such effect of HPV infections that is characteristic and considered pathognomic of it, is Koilocytosis or as it was first described Koilocytic atypia, which has been suggested to reflect the HPV load in the lesion. It was through the identification of these koilocytes in 35% of Oral Squamous cell carcinoma cases that in 1983 Syrjanen et al put forward their probable role in oral carcinogenesis. They also observed similar changes in oral potentially malignant disorders further suggestive of their oncogenic potential. Further evidence to this thought was provided in 2007 by D'Souza et al who showed a positive correlation between HPV and oropharyngeal lesions.1, 2, 3
Koilocytes:
History:
With the advent of the PAP smear in 1941 for the diagnosis of cervical lesions numerous cytological observations were made over a short period of time. In 1951, Canadian gynaecologist/ cytologist Dr J. Earnest Ayre described and illustrated halo cells while working in Miami. He described these squamous cells of the uterine cervix to be having a perinuclear clearing, mono or bi-nucleate with poor keratinizing features and hyperchromatic atypical nuclei. He also proposed that these vacuolated cells were “pre-cancerous” and being always found in association with chronic inflammatory cells, must be caused by a long standing inflammation of infection either viral or otherwise. He said that the presence of these vacuoles denoted a degenerative change of the cell.3
In a letter of correspondence to the Wiley online library celebrating the 57th birthday of koilocytes, Leopold G. Koss recounts on how he and his associate technician Grace Durfee came across these cells. In 1955 George Papanicolaou, a three times visitor to their laboratory noticed the cells in cervical smears and labelled them 'dyskaryotic', but failed to realise the potential of their unusual appearance. Koss and Durfee, fascinated by these cells, further described them in a series of lectures on cytopathology in Europe in the year 1955.4
As he studied the cells the feature that struck him the most was their hollow appearance and he related this largely to a term used to describe hollow nails- Koilonychias and hence the name Koilocytosis. In 1956 they published their findings in a study titled “Unusual patterns of squamous epithelium of the uterine cervix: cytological and pathologic study of Koilocytic atypia” where koilocytes were described as- 'large epithelial cells with relatively small but irregularly shaped hyperchromatic nuclei surrounded by glycogen negative clear halos'. Subsequently zur Hansen associated koilocytes to the Human Papilloma virus in his article “Human papillomaviruses and their possible role in squamous cell carcinomas” in the year 1977 for which he received the Nobel Prize in 2008.4
Light microscopy:
Koilocytes may be defined as “Squamous epithelial cells having hyperchromatic nuclei with irregular outlines that are displaced eccentrically by a large perinuclear halo that may take up almost the entire cytoplasm and show occasional artefactual multi-nucleation” Due to their characteristic appearance they have been described as – “Halo cells that are no angels”. These cells have been known to show the presence of 50 to 100 viral particles on ultrastructural observation.
Features of koilocytes:
Dyskeratosis and cells rich in keratohyaline granules are residual effects of viral replication and do not correlate with the viral load.1,2,5
Koilocytosis:
The process of formation of these characteristic cells in HPV infections where the pathogenetic process primarily involves the nucleus and not the cytoplasm, has long remained a mystery and in ways still does. Many authors have suggested that these cell represent forms of apoptosis and senescence but with no substantiating evidence. Koilocytosis in essence can be explained if the mechanism of generation of perinuclear vacuole comes to light. Krawczyk E and co-workers in 2008 did precisely this, explaining the role of the HPV oncoproteins in influencing the formation of the perinuclear vacuole. They chose to work with the transforming oncoproteins of high risk HPV- 16: E6, E7 and E5. Among the three oncoproteins it is well understood that E6 and E7 encode for proteins that act on the p53 and Retinoblastoma genes immortalizing the cell, however the role of E5 is poorly understood and numerous studies are and have been conducted regarding its pathogenic role in HPV induced lesions and carcinomas.5
They utilized vectors carrying retroviruses encoded for 16E6 and 16E7 proteins to immortalize human ectocervical cells, and following which 16E5 encoding cells were liberated using 16E5 retroviral vectors. E6 and E7 encoding cells were also produced similarly. Human foreskin keratinocytes were also encoded for HPV16 E6 and/or E7. The cells were fixed, stained with Haematoxylin and Eosin stains and visualized. They observed perinuclear vacuolisation similar to koilocytes seen in cervical PAP smears in immortalized E5 expressing Human ectocervical cells with few qualitative differences such as multi-loculation of the perinuclear vacuole. In a further attempt to relate Koilocytosis to the E5 protein they used E6 and 7 immortalized cells with and without E5 expression and found that koilocytes were readily detectable in the E5 expressing group. Immunohistochemical staining of these cells with an antibody toward the epitope tagged to the E5 protein showed presence of E5 on the Koilocytic vacuolar membranes and in the cytoplasm. He further went to show that E5 did not act alone in the process, was unable to generate koilocytes by its lone expression and did so in conjunction with E6. He noted that co-expression of E7 drastically reduced koilocyte formation suggesting a suppressive role. The active region of E5 involved in Koilocytosis was proposed to be present among the twenty terminal amino-acids of E5, as mutants with deletion of ten terminal amino-acids showed nearly 50% reduction in Koilocytosis and no Koilocytosis was seen in those with deletion of twenty terminal amino-acids. The property of multi-vesiculated perinuclear clearing was seen to be more in the former mutant and was ascribed to incomplete vesicle fusion. They said that the lack of Koilocytosis was not due to reduced expression of these proteins, but rather a result of altered activity. Similar results were obtained with low risk HPV-6 and in Human Foreskin cells. The authors say the reason for small differences in Koilocytosis in-vitro and those seen in the original disease process may be because the levels of expression of E5 in-vitro may not be as high as that in a productive infection, the fact that the cells the studied were not terminally differentiating and that only specific viral oncogenes were employed, not the entire genome that may possess additionally required genes. Even under conditions that acutely induce cell differentiation (Ca2+ and 10% foetal bovine serum) there was no increase in Koilocytosis in-vitro suggesting that the oncogenes may require more stringent conditions of differentiation for their expression by the respective retroviral promoters. The production of koilocytes by E5 is supported by the fact that the expression of E5 protein is increased in the upper mature layers of the epithelium where Koilocytosis occurs.5
In another study Krawczyk shows that E5 binds to the perinuclear cytoplasmic protein Karyopherin beta 3 belonging to the nuclear import receptor superfamily. It is seen in the host cell cytoplasm and in the perinuclear area bound to some nucleoporins. These proteins regulate the nuclear import of various material such as- ribosomal proteins, histones and in case of infection- viral proteins such as the L2 of HPV-16 and 11. They may also act as cytoplasmic chaperones and microtubule motor adaptors and are found localized in the perinuclear cytoplasm. In a similar way to their previous study cells were infected with 16E5 positive viral vectors with and without the C-terminal 10 amino acids and studied. Cojoined expression of E5 and Karyopherin were seen in the perinuclear region along with the observation that E5 protein was seen sandwiched in between the nucleus and cytoplasmic karyopherin expression in Human ectocervical cells. They also showed that the binding of the E5 protein to Karyopherin occurred in the realms of the C-terminal 10 amino acids as the amino acid deletion mutant showed largely defective bonding with Karyopherin. The role of this interaction between the two is not evident though the authors correlate the function of karyopherin as a vesicle transporter ad the known ability of E5 to prevent vesicle transport and suggest a role involving this. An SiRNA mediated knockout and down regulation of Karyopherin expression also caused a decrease in Koilocytosis and thus placed Karyopherin as one of the important Koilocytosis mediating proteins.6
Krawczyk et al also use markers for cellular senescence Beta galactosidase and show that being detectable in only a very small number and show that koilocytes probably do not represent process of apoptosis or senescence.5
MULTINUCLEATION or multilobulation?
Koilocytes apart from possessing a perinuclear halo also may present with multinucleation. Current research has shown that what appears as multinucleation is in fact an artefactual appearance of a multilobulated nucleus. Cho NH et al in 2005 and 2006 used 3-dimensionally reconstructed confocal fluorescent images of cervical swab and tumor samples with koilocytes to better inspect the cell. They showed that what was seen in 2-dimension as multinucleation was in fact multilobulation of a single nucleus, with several lobes joined together by an anaphase bridge. They attributed this anomaly to be a result of the oncoproteins inducing chromosomal instability and abnormal p53 expression, cumulative with G2 cell cycle checkpoint aberrations that cause a non-dividing nucleus that is hence multi-lobulated. The multilobulation may also be thought to coincide with an increased viral load, but further studies may be required to confirm this assumption.7, 8
E5 protein:
The E5 protein is a small hydrophobic protein of the HPV16 with 83 amino acids that is localised on the membranes of endoplasmic reticulum and nuclear envelope.9 The functioning of E6 and E7 proteins in HOV pathogenesis is a fairly well understood phenomenon. But the activity of E5 in HPV is vast and the understanding of its different roles in pathogenicity is growing still. 5No intrinsic enzyme activity is possessed by this protein and it is thought to effect oncogenic transformation through interactions with several cellular proteins and cell wall lipid rafts.(6) It has been implicated in
Disbrow G L and co-workers in 2003 showed that E5 proteins of HPV-16 have the ability to limit cell proliferation and reduce the life span of keratinocytes. Condjella et al in 2009 demonstrated similar property of Canine papilloma virus E5 protein and also noted the increase in cells in the G1 phase, reduction of cells in the S phase and induction of Endoplasmic Reticulum stress. They also noted that these effects were considerably reduced in the presence of E6 and E7 genes. 10Though specific proteins have been pointed out that seem to serve as targets for E5 the varied propertied of E5 tend to suggest that there can be several targets through which it effects its actions. 5Though known to be localised on the membranes of endoplasmic reticulum the orientation and active domain of the protein came to light recently in an experimental design by Krawczyk and co-workers in 2010 before which the understanding of orientation was greatly through hydrophobicity plots. They employed immunofluorescence with differential detergent permeabilization technique tagging both ends and identified that the C-terminal of the protein is oriented toward and exposed to the cytoplasm. They also deleted the third trans-membrane domain of the protein and showed that this resulted in the C-terminal end being restricted within the endoplasmic reticulum lumen thus suggesting a 3 pass transmembrane topography of the protein.9 The study results are in accordance with their previous assumptions while working with Karyopherin- that since karyopherin binds with the C-terminal and is localized in the cytoplasm, the C-terminal end of E5 must be exposed to the cytoplasm also.(6,9) Their study also negates assumptions by other authors who suggest that E5 is localized on the plasma membrane of the cells with C-terminal exposed extracellularly aiding in cell to cell fusion and no immunofluorescence localization of E5 was seen in the plasma membrane.9
TRUE AND ABORTIVE KOILOCYTES:
Lhee MJ in 2014 proposed a concept of true and abortive koilocytes and their use in a system for prognostication or infectious fate of the HPV infection. The criteria for defining either of them are:11
True Koilocytes:
Abortive Koilocytes: Cells that have lost the capacity to replicate virions
Fixative for koilocytes
The origin and existence of koilocytes has been shrouded for long and through newer methods of continuing research is slowly coming to light. At this point it is worthy to note that all that is possible to be evaluated at a light microscopic level does not provide as much information as more advanced methods do. This may in part be due to the fact that the koilocyte is for all means- a clear cell. When posed with similar situations in the past research advancements in understanding the cell has in part focussed on what component of the cell may be lost in giving it this clearing and what could be causing the loss of that material. As in the case of myelin and lipid the process of tissue preparation for light microscopic study itself was the cause. Possibly along the same line of thought Patil S and co-worker in 2015 set out to determine what fixative could best bring out the features of these cells at a light microscopic level. They utilized three common fixatives formalin, Osmium tetra-oxide and glutaraldehyde and evaluated which best expressed koilocytes. They found that Osmium tetra-oxide provided the best results with sharp and clear delineation of cytoplasmic and nuclear details. As for the other two formalin was seen to be superior to glutaraldehyde.12
Subscribe now for latest articles, news.